|

6.9. Endemismos
Los endemismos
son taxones (pueden ser especies, géneros, familias...)
o comunidades vegetales y animales, cuya "área es significativamente
inferior al área media de los taxones de su mismo rango" (Costa, 1998).
No siempre son especies con un área de distribución pequeña, como la
Salsola
genistoides que habita una pequeña región del litorial
murciano
y almeriense, al sureste de la península Ibérica (Alcaráz Ariza, 2008).
También el orden de los monotremas (Monotremata),
compuestos por los
ornitorricos y los equidnas, es considerado endémico de la amplia
región Australiana.
Los endemismos, tienen un gran valor biogeográfico, y por ello se han
empleao habitualmente en la caracterización de los territorios (Costa,
1998). De hecho, en las clasificaciones de los reinos y regiones
biogeográficas del planeta (ver capítulo
de reinos biogeográficos) se
tienen muy en cuenta el número y tipo de endemismos de cada zona.
En la formación de un
endemismo intervienen muchos factores históricos
(y paleontológicos), ecológicos, fisiológicos, antropológicos...
Influyen, por lo tanto, elementos propios de la especie (como por
ejemplo que esté adaptada a un rango térmico bajo o que tenga una baja
tasa de dispersión de las diásporas) y elementos externos (bióticos,
como la aparición de una especie competidora; o abióticos, como la
formación de un sistema montañoso). Aunque la causa principal suele ser
una reducción de su área de distribución o que el taxón haya quedado
aislado.
Los endemismos pueden
quedar aislados por las siguientes causas (o por
la combinación de varias):
- Por un aislamiento
geográfico, es una de las causas más comunes, por
ello las montañas, las islas, los desiertos y las áreas con suelos
especiales, suelen ser territorios con unas altas tasas de endemismos,
y con una flora y una fauna muy original.
El endemismo montano
(u orofitismo) es clave, aunque depende de muchos
factores, y suele aumentar en función de la altura de la montaña y del
aislamiento geográfico respecto a otros relieves. Las montañas suelen
poseer muchos endemismos debido a que tienen un clima y una orografía
muy peculiar (con suelos delgados, terrenos escarpados, peligro de
aludes, inaccesibilidad, formación de microclimas...).
El porcentaje de endemismos puede ser muy variable, pero tiende a ser
alto. La flora orófila endémica de los Apeninos (Italia) es del 13 %,
de los Alpes Centrales (Italia, Francia, Suiza) es del 18%, en Pirineos
supone un 14%, mientras que en sierra Nevada (España) es del 36% y en
las montañas Corsas (Grecia) del 38% (Favarger, 1972). Pero en el monte
Kenia (en Kenia) la cifra asciende a más del 80%, debido a que es una
montaña-isla (en sentido biogeográfico), que se encuentra aislada, ya
que no forma parte de ninguna cadena montañosa (Favarger, 1972).
De hecho, prácticamente todos los macizos montañosos del mundo poseen
una flora y una fauna endémica que les caracteriza y que les diferencia
de los demás, a pesar de tener incluso un origen ancestral común. Por
ejemplo, hay numerosas especies endémicas distintas del género
Gentiana,
en los Alpes, el Himalaya y en las montañas de China; y
dentro de la familia de las Gesneriáceas,
la especie Ramonda
myconi es
endémica de los Pirineos, mientras que las especies Ramonda nathaliae y
Ramonda
serbica son únicas de los Balcanes.
Otros ejemplos de endemismos montanos muy característicos son, a nivel
de plantas la Saxifraga
florulenta situada en el parque nacional de
Mercantour (Francia) o el Thymus
godayanus que se localiza en las
montañas más altas de la Comunidad Valenciana (España), y a nivel de
animales, el majestuoso cóndor (Vultur
gryphus) en los Andes o el
tritón pirenaico (Calotriton
asper) que se distribuye por los Pirineos
(en España, Francia y Andorra) y por algunas montañas de Cataluña como
el Montsec, la sierra de las Guillerías y el Montseny.
El endemismo insular
(o insularismo) tiene una gran importancia,
sobretodo en islas con grandes altitudes, como en Córcega; o en las
islas volcánicas formadas recientemente y alejadas del continente, como
las islas Canarias que surgieron en el Mioceno y que actualmente tienen
unas 460 especies endémicas, de las 1.400 que componen la flora
canaria, y unos 20 géneros endémicos, de los 330 que componen la flora
canaria (Costa, 1998). Y en cuanto a fauna, de los 6.843 artrópodos
presentes en las islas Canarias 2.704 son endémicos; de los 5.668
moluscos, son
endémicos 2.172; y de los 123 cordados son únicos de las islas 22, 14
reptiles, 5 aves y 3 mamíferos (Esquivel et al, 2001).
|
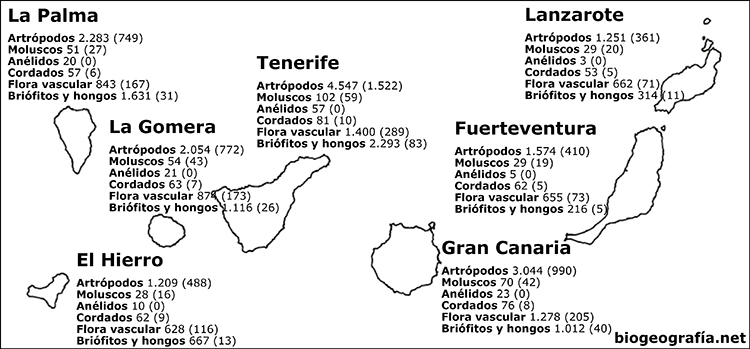 |
Mapa de endemismos de
las islas Canarias.
Número de especies de cada grupo (Número de endemismos canarios).
Esquivel et al, 2001. Elaborado por Alberto Díaz, 2019. |
En las islas, la tasa de endemismos de flora y fauna terrestre suele
ser muy alta, puesto que el mar (o el océano) actúa como barrera y
elemento aislante. En las islas Baleares (España) hay un 7% de plantas
endemicas, 10% en Creta (Grecia) y un 30% en Córcega (Francia). Pero
hay islas con porcentaje mucho mayores. En Madagascar los endemismos
suponen un 65 % de la flora, en Nueva Zelanda existen alrededor de
1.000 especies vegetales endémicas, más del 72-80 % del total (Lacoste,
1973; Sanchis et al, 2004). Y en las islas del Pacífico el porcentaje
de endemismos en muchos archipiélagos es enorme, en Hawai debido a su
aislamiento geográfico y a su peculiar origen posee un 82-92% de
endemismos, y en Nuevas Hébridas y Nueva Caledonia los
endemismos llegan casi al 100% (Costa, 1998).
En animales podemos destacar a gran parte de la fauna australiana, o la
isla de Ceilán en donde más de la mitad de los reptiles y mamíferos son
exclusivos de esa isla. O la isla de Madagascar en donde son endémicos
todos los anfibios, el 90 % de los reptiles (la mitad de las especies
de camaleones del planeta), el 55 % de sus mamíferos (como los lémures
y los fosas) y el 50 % de sus aves (con cinco familias enteras
endémicas: Bernieridae,
Brachypteraciidae,
Leptosomidae,
Mesitornithidae
y Vangidae)
(IOC, 2016).
Aunque también hay islas con niveles muy bajos de endemismos como las
islas Británicas (Ferreras, 1999). En Gran Bretaña la acción del
ser humano, la uniformidad del clima y el hecho de que aún estuviera
unida al continente europeo después de la última glaciación, hicieron
que las islas sean pobres en especies endémicas animales y vegetales,
salvo en las montañas de Gales y Escocia (Costa, 1998).
El endemismo desértico
es la consecuencia de las especiales
características de los desiertos. Estas áreas del planeta aparte de
actuar como barreras naturales (que las independiza de otros entornos),
también generan un ambiente particular que hace que las especies que se
han adaptado a vivir allí presenten unos fenómenos de convergencia
adaptativa muy característicos, y una mayor competencia frente a las
especies que pretenden invadirlas.
En los desiertos la tasa de endemismos suele ser alta, a pesar de que
haya pocas especies que puedan vivir en ellos. En el desierto del
Sáhara se calcula que el 25% de las plantas son endémicas, y en el
desierto de Sonora, la tasa de endemismo es del 26%, pero en las
montañas que se encuentran dentro de estos desiertos la tasa es mucho
mayor. Por ejemplo, dentro del desierto del Sáhara, en la meseta de
Tassili la cifra aumenta al 40%, y en el macizo de Ahaggar o de Hoggar,
la flora endémica representa el 60% del total de especies.
El endemismo edáfico o
edafismo, se forma por la adaptación de las
plantas y los animales a suelos y sustratos característicos. En
terrenos con suelos salinos, poco fértiles, con metales pesados y
sustancias tóxicas (como cobre o mercurio), en áreas yesíferas, en
sitios con silicatos básicos (dolomías, serpentinas...) y rocas
calaminares (ricas en zinc), campos de dunas... suele haber una gran
variedad de endemismos. Estos lugares con suelos especiales representan
espacios aislados del entorno, y en ellos suelen vivir taxones muy
especializados.
Como ejemplos de endemismos edáficos destacan las plantas
serpentinícolas Cupressus
sargentii de California (Estados Unidos),
Silene
pindicola y Arenaria
serpentini de las cumbres del Pindo y del
Olimpo (Grecia), o Digitalis
laciniata y Teucrium
reverchoni de las
sierras Bermeja, Carratraca y del Agua (en el sur de España); plantas
adaptadas a suelos dolomíticos como Anthyllis tejedensis,
endémica de
las sierras Béticas (también en el sur de España); y plantas endémicas
y gipsófilas como Gypsophila
struthium, Helianthemum
squamatum,
Lepidium
subulatum, Ononis
tridentata... (Costa, 1998).
- Por un aislamiento
genético, que suele estar ligado a un aislamiento
geográfico, puesto que es debido a una interrupción de la comunicación
o relación de la especie o el taxón con sus comunidades vecinas.
Generalmente, al producirse el aislamiento genético, los individuos que
han quedado aislados deben reproducirse entre ellos, de manera
endogámica, lo cual provoca cambios morfológicos (tras varias
generaciones) o fenómenos de esterilidad que pueden hacer que la
especie desaparezca.
- Por un cambio brusco
de las condiciones del medio, el aumento de la
aridez, las glaciaciones, las variaciones de temperatura y de
humedad... puede generar cambios en las especies por adaptación a las
nuevas condiciones o puede provocar las fragmentación de las áreas de
distribución de las especies.
Dado a la gran cantidad de tipos
de endemismos, éstos se suelen
clasificar en función de su rango taxonómico, de su antigüedad, del
territorio que ocupan y de su origen.
Los endemismos se dividen en
función de su rango taxonómico en:
- macroendemismos
o megaendemismo
que se refieren a rangos superiores
al de especie (géneros, familias, órdenes....), y suelen ser debidos a
los movimientos de las placas tectónicas, como la subclase de los
Prototheria,
compuesta por ornitorrincos y equidnas, situados en la
región de Australia.
- microendemismos,
generalmente de origen más reciente, y referidos a
especies, subespecies o variedades, como el Silene diclinis, un
endemismo que se localiza únicamente en las proximidades de la
localidad de Xátiva; o el Limonium
mansanetianum que sólo se encuentra
en una pequeña área de Valencia (Sanchis et al, 2004).
Los endemismos también se clasifican en función de su antigüedad
(según
Engler, 1882) en:
- paleoendemismos
o endemismo relictos,
son taxones
antiguos, surgidos
durante el Paleozoico y el Mesozoico. Representan especies aisladas
filogeneticamente por la extinción de sus parientes más cercanos, y que
frecuentemente tienen un alto nivel de poliploidia.
Muchos paleoendemismos, son endemismos
residuales, es decir, en el pasado
geológico estos taxones tenían un área de distribución mucho mayor al
actual y estaban mucho más diversificados, pero hoy en día suelen ser
las únicas especies de su grupo taxonómico y ocupan un espacio muy
limitado (Costa, 1998; Mota et al, 2003).
Los paleoendismos más característicos, a nivel de plantas, son los
ginkgos (Ginkgo biloba),
únicos supervivientes de la clase
Ginkgoopsida,
que aparecieron en el Pérmico y tuvieron su mayor auge en
el Jurásico (cuando unos 20 géneros y más de 100 especies de ginkgos
ocupaban gran parte del hemisferio norte), pero que actualmente sólo
viven de manera natural en las montañas del este de China (aunque se
cultivan artificialmente en muchos jardines de Europa y América).
También son paleoendemismos los bosques de secoyas gigantes
(Sequoiadendron giganteum)
y secoyas rojas (Sequoia
sempervirens) de
California (Estados Unidos); las metasequoias (Metasequoia
glyptostroboides) de las provincias chinas de Sichuan y
Hubei; la
familia de las podocarpáceas (Podocarpaceae)
de Chile, África y Nueva
Zelanda; las araucarias (género Araucaria)
de Sudamérica y
Nueva Caledonia; las welwitschias (Welwitschia
mirabilis) del desierto
del Namib, o Idria
columnaris y Fouqueria
splendens del desierto de
Sonora (Costa, 1998).
- y los neoendemismos,
que son los taxones con un origen más reciente,
formados a veces, después de las glaciaciones. De hecho, los procesos
de formación de los neoendemismos siguen actuando en la actualidad.
Muchos
neoendemismos o endemismos recientes, se forman como resultado de la
especiación en medios aislados Mangenot (1972). Y muchos de ellos,
aparecen en territorios con una historia geológica muy reciente (como
islas volcánicas, desiertos...).
Como se puede observar en la siguente tabla, diversos autores han
empleado conceptos sinóminos o muy parecidos, para referirse a los
paleoendemismos y los neoendemismos.
| Autor |
Endemismos antiguos |
Endemismos recientes |
| Engler (1882) |
Paleoendemismo |
Neoendemismo
|
| Drude (1890) |
Endemismo relictual |
Endemismo secundario |
| Briquet (1901) |
Endemismo por conservación |
Endemismo por innovación |
| Braun-Blanquet (1929) |
Endemismo paleógeno |
Endemismo neógeno |
| Ridley (1925) |
Epibiótico |
Endémico
|
| Herzog (1926) |
Endemismo antiguo |
Neoendemismo |
| Wherry (1944) |
Endemismo secundario |
Endemismo primario |
Favarger y
Contandriopoulos (1961) |
Paleoendemismo,
patroendemismo, y
esquizoendemismo antiguo |
Apoendemismo y
esquizoendemismo incoativo |
| Mangenot (1972) |
Endemismo pasivo |
Endemismo activo |
| Conceptos
sinóminos de paleoendemismos y los neoendemismos. Mota et al, 2003;
Alberto
Díaz,
2020. |
En
función
de su origen,
y teniendo en cuenta criterios sintaxonómicos, los botánicos Claude
Favarger y Juliette Contandriopoulos (1961) clasificaron los endemismos
en:
- esquizoendemismos
son endemismos emparentados que se han formado a
partir de un ancestro común, endémico o no, cuya área ha sido
fragmentada y cuyas poblaciones siguen una diferenciacion gradual,
lenta,
progresiva, y simultánea en cada una de sus áreas (Costa, 1998; Mota et
al, 2003). Los
esquizoendemismos son endemovicariantes entre sí, y tienden a ocupar
áreas disyuntas. Presentan características cromosómicas muy
parecidas (poseen el mismo número de cromosomas), y morfológicamente
son
todos muy similares.
Muchos endemismos de origen montano e insular son esquizoendemismos.
Por ejemplo, el género Hippocrepis
(plantas de la familia
Fabaceae)
presenta diferentes especies muy similares (puesto
que tienen un origen muy reciente), pero con poblaciones
diferenciadas en un territorio relativamente pequeño situado al este de
la península Ibérica. La especie Hippocrepis
valentinase se encuentra
en la Comunidad Valenciana, entre las provincias de Alicante
y Valencia; Hippocrepis
balearica en la isla de Mallorca e
Hippocrepis
grosii en Ibiza.
Además,
es importante tener en cuenta que los esquizoendemismos se forman
independientemente de la antigüedad de la línea de parentesco (Mota et
al, 2003). Existen esquizoendemimos antiguos o relictos, del Terciario,
como Ramonda myconi del Pirineo (y R. nathaliae y R. serbica, de los
Balcanes) o Viola cazorlensis (y V. kosanini de Montenegro y V.
delphinianta del Monte Olimpo); y esquizoendemimos recientes,
postpleistocénicos, como Petrocoptis o Andryala agardhii y A. maroccana
(Blanca y Valle, 1986).
- paleoendemismos
son "taxones aislados desde el punto de vista sistemático" (Mota et al,
2003). Pertenecen a géneros monotípicos, con una sola especie.
Los
paleoendemismos no han tenido que originarse necesariamente en su área
actual, y su distribución moderna suele ser una reliquia de un
territorio antiguamente más amplio. Con frecuencia son diploides, pero
no siempre.
En España destacan como paleoendemismos del Terciario la hierba de
Rothmaler (Rothmaleria
granatensis) y el pinsapo (Abies pinsapo)
(Blanca y Valle, 1986).
- patroendemismos
son taxones primitivos de carácter endémico y diploides que
dan
lugar a la formación, en las proximidades de su área, a taxones
correspondientes con mayor nivel de ploidía (poliploides), y que son
llamados
taxones de sustitución (Costa, 1998; Mota et al, 2003). Suelen
colonizar
rápidamente grandes extensiones, y tienden a tener un área de
distribución mayor que la del taxón patroendémico inicial (diploide).
Un ejemplo de patroendemismo es el de la pequeña planta andaluza Crepis granatensis
(Blanca y Valle, 1986).
- apoendemismos
son el caso contrario de los patroendemismos, es decir, "endemismos de
carácter poliploide que tienen su
origen en un territorio determinado por poliploidización a partir de un
taxón diploide más o menos extendido en los territorios vecinos"
(Costa, 1998). Es decir, los taxones derivan unos de otros, y los más
recientes ocupan áreas más pequeñas que los más antiguos.
Cuando
el ancestro (antecesor o precursor) diploide de un taxón apoendémico no
se conoce o ha desaparecido, el endemismo cae en la categoría del
esquizoendemismo (Mota et al, 2003).
Como ejemplo de
apoendemismo ibérico destaca la silicícola Centaurea citricolor que es
probablemente un autopoliploide de la calcícola Centaurea monticola
(Blanca y Valle, 2003).
- criptoendemismos
son taxones que tienen un gran potencial endémico pero que
aún no han sido descritos. Generalmente, su número cromosómico es
diferente al del taxón original (cuya distribuición es mucho más
amplia), y son tan distintos morfológicamente que es imposible
distinguirlos (Mota et al, 2003).
Por último, en función
del
territorio que ocupan las especies endémicas se dividen
en:
- semiendémicas:
en el caso de especies que sólo son endémicas en su
período de migración.
- cuasiendémicas:
cuando ocupan un área pequeña de distribución, pero
también tienen pequeñas poblaciones marginales fuera de ese área. |
|
Continúa en la siguiente
página con el apartado
de vicarianza...
|

| |
 |
|
 |
|
www.biogeografia.net
|